3 註解
吸蟲(trematode)屬扁形動物門(Platyhelminthes)的吸蟲綱 (Trematoda)。該綱下隸3個目,即單殖目(Monogenea)、盾腹目(Aspidogastrea)和複殖目(Digenea)。單殖目吸蟲爲水生冷血脊椎動物特別是魚類的寄生蟲。盾腹目吸蟲爲軟體動物、魚類和龜、鱉等的寄生蟲。寄生人體的吸蟲都屬複殖目。複殖目的吸蟲種類繁多,生活史複雜、具有有性世代和無性世代交替,無性世代在軟體動物中寄生,有性世代大多在脊椎動物體內寄生。
4 吸蟲的形態
大多數複殖目吸蟲成蟲身體扁平、呈葉狀或舌狀,少數呈扁錐形或近圓柱形,大小依蟲種而異。蟲體表面爲光滑或具有小棘的角質層覆蓋。吸盤爲附着器官,通常有2個,其中一個包圍着口孔,稱口吸盤,另一個位於腹面,稱腹吸盤。生殖孔通常位於腹吸盤的前緣或後緣處,個別蟲種具生殖吸盤。排泄孔位於蟲體的末後端。
4.1 體壁
吸蟲的體壁是由外層的皮層(tegument)和皮層下的細胞體構成,系合胞體(syncytium)結構,覆蓋於蟲體的體表。皮層和細胞體之間有胞質小管相通。皮層整層爲胞質性,無核也無細胞界線,由外質膜(external plasma membrane)、基質(matrix)和基質膜(basal plasma membrane)組成。感覺器位於基質中,有纖毛伸出體表之外,另一端有神經突(nerve process)與神經系統相聯。基質膜之下爲基層(basement layer), 基層之下爲外環肌和內縱肌。 皮層細胞(tegumentary cell)位於肌層下,很大,內有胞核、內質網、核糖體(ribosome)、吞噬體(phagosome)、線粒體和高爾基體。有許多胞質通道與基質相通,有的甚至通到實質細胞(parenchymal cell),胞質內及胞質通道中均有許多分泌小體。吸蟲的體壁具有保護、吸收營養和感覺等功能。

4.2 消化系統
包括口、前咽(prepharynx)、咽(pharynx)、 食道及腸管。口由口吸盤圍繞,位於蟲體的前端或偏腹面。前咽短小或缺如。咽爲肌質構造,呈球狀。咽和腸管之間爲細長的食道,食道的兩側常有若干個單細胞腺體,各有管道通向蟲體前端。腸管分左右兩支向蟲體後端延伸,絕大多數蟲種的兩條腸管在蟲體後端形成封閉的盲端,不再合攏,少數吸蟲(如裂體科)的兩條腸管在體後部聯合成單一的盲管。吸蟲無肛門,未被消化吸收的廢物經口排出體外。
4.3 排泄系統
由焰細胞(flame cell)、毛細管、集合管、排泄囊、排泄管和排泄孔組成。焰細胞爲凹形細胞,具有一個大的細胞核,顯微鏡下核仁明顯可見,在凹入處有一束纖毛。焰細胞因其纖毛顫動時似火焰跳躍而得名。排泄液借纖毛的顫動而進入胞腔,然後經毛細管、集合管集中到排泄囊,最後從排泄孔排出體外。複殖目吸蟲的排泄孔只有一個, 位於蟲體的後端。

4.4 神經系統
復殖吸蟲的神經系統不發達。在咽的兩側各有一個神經節,相當於神經中樞。神經節間彼此有背索相連。兩個神經節各發出前後三條神經幹,分佈於背面、腹面及側面。向後伸展的神經幹,在幾個不同的水平上皆有橫索相聯。感覺末梢由前後神經幹發出到達口吸盤、咽、腹吸盤等器官,以及體壁外層中的許多感覺器。
4.5 生殖系統
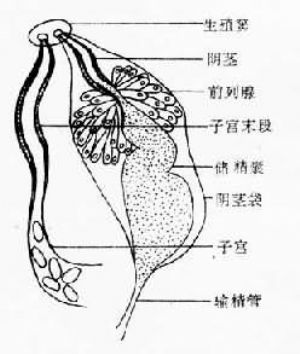
復殖吸蟲中除了裂體科是雌雄異體外,其它均爲雌雄同體。雌雄生殖孔開口於生殖竇(genital sinus)內,或各自由生殖孔開口。
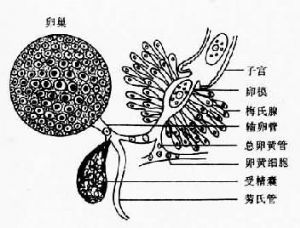
雄性生殖器官包括睾丸、輸出管、輸精管、貯精囊、射精管、陰莖袋、前列腺與陰莖。陰莖開口於生殖竇或生殖孔,交配時陰莖可經生殖孔伸出體外與雌性生殖器官的遠端相交接。雌性生殖器官包括卵巢、輸卵管、梅氏腺(Mehlis’s gland)、卵模(ootype)、卵黃腺(vitelline gland)及子宮等。另外還有勞氏管(Laurer’s canal),其一端接受精囊或輸卵管,另一端向背面開口或成爲盲管。 卵黃腺由許多卵黃泡組成。卵黃細胞或卵黃球由卵黃泡經小管、上下縱管、橫卵黃管、卵黃囊進入卵黃總管。卵黃總管與輸卵管匯合後在子宮的起點前,其周圍有一羣單細胞腺即梅氏腺,被梅氏腺包圍的部分稱卵模。卵細胞由卵巢排出後在輸卵管中與精子相遇而受精,然後與由卵黃管排出的卵黃球一同進入卵模。各蟲種卵殼的特殊形狀爲卵模收縮的結果。卵逐個由卵模推進子宮管,並不斷向遠端移動,卵的成熟程度隨移動而增加。子宮長短不一,靠近生殖孔的一段稱爲子宮遠端(metraterm),爲肌質結構,蟲卵由此排出;子宮遠端尚有陰道的作用,吸蟲可進行異體受精或自體受精。
5 吸蟲的生活史
復殖吸蟲的生活史複雜,不但具有世代的交替(含有性世代和無性世代),還有宿主的轉換。宿主的轉換包括有性世代寄生的宿主(終宿主)和無性世代寄生的宿主(中間宿主)的轉換,有些吸蟲在無性世代還需轉換宿主(第一中間宿主、第二中間宿主等)或通過轉續宿主進入終宿主體內。復殖吸蟲的第一中間宿主爲淡水螺類或軟體動物,第二中間宿主依蟲種不同可爲魚類、甲殼類或節肢動物等。終宿主大多爲脊椎動物和人。
復殖吸蟲的生活史離不開水,蟲卵必需入水或在水中被軟體動物吞食後才能孵化出毛蚴(miracidium),毛蚴進入中間宿主後發育爲胞蚴(sporocyst),胞蚴體內的胚細胞經反覆分裂後分化成許多雷蚴(redia),最後從母體逸出。胞蚴和雷蚴都可以不止一代,有的蟲種可繼續產生三、四代雷蚴,每代雷蚴都同時含有尾蚴(cercaria)及下一代的雷蚴。胞蚴或雷蚴中的尾蚴成熟後,在一定的外界條件影響下即可從母體逸出,藉助尾部的擺動,在水中游動,在某些物體上結囊形成囊蚴(metacercaria),或在第二中間宿主體內發育成囊蚴。裂體科的吸蟲無囊蚴期,尾蚴直接侵入終宿主經童蟲發育爲成蟲。囊蚴進入終宿主消化道後,後尾蚴即脫囊而出變爲童蟲,在適宜的寄生部位發育爲成蟲。
6 吸蟲的生理
寄生吸蟲在其種系發生過程中不僅在形態、結構上發生了一系列與寄生生活相適應的變化,而且也產生了適應寄生生活需要的生理功能。吸蟲的營養來源主要爲腸內容物、腸粘膜、血液或組織液,具體營養來源依蟲種和寄生部位而定。其消化過程主要在腸內進行,但有的吸蟲兼有細胞外和細胞內消化。寄生吸蟲與宿主體液間有一層變動着的界面,這種界面既存在於蟲體的體表,也存在於蟲體消化管道的內面。氣體(如氧)和小分子物質可經此界面直接進入蟲體,如有的吸蟲可以通過體表皮層吸收葡萄糖、氨基酸、維生素、核苷等。因此,此層界面可稱爲營養界面。吸蟲的代謝產物、分泌物等也可經此界面排出體外。
吸蟲獲得能量的方式主要爲有氧代謝和無氧代謝。葡萄糖和糖原等碳水化合物被認爲是吸蟲重要的能量來源。大多數寄生吸蟲的成蟲主要依靠糖的無氧酵解獲得能量,但在某些吸蟲的幼蟲期,還需要從有氧代謝中獲得一定的能量,以滿足快速生長的需要。
吸蟲生活史過程中基本是需氧的,但吸蟲缺呼吸系統,氧主要是由體表、消化道內壁或其它與氧接觸的部位進入體內,有的蟲種氧是由食物攝入到消化道,然後再向組織擴散。由於不同吸蟲寄生的部位不同,因此所處環境的氧含量差別可以很大。即使同一種吸蟲,不同發育階段的蟲體對氧的需求也不一樣。因此,長期適應的結果使得吸蟲獲得了良好的調節氧的消耗率的能力,並能在氧分壓低時更經濟地利用氧。
7 吸蟲的分類
科 | 屬 | 種 | 寄生部位 |
後睾科 Opisthorchiidae | 支睾屬 Clonorchis | C. sinensis | 肝膽管 |
異形科 Heterophyidae | 異形屬 Heterophyes | 異形異形吸蟲 H. heterophyes | 腸管 |
片形科 Fasciolidae | 薑片屬 Fasciolopsis | F. buski | |
片形屬 Fasciola | F. hepatica | 肝膽管 | |
並殖科 Paragonimidae | 並殖屬 Paragonimus | P. westermani | 肺 |
狸殖屬 Pagumogonimus | P. skrjabini | ||
裂體科 Schistosomatidae | 裂體屬 Schistosoma | 日本裂體吸蟲 S. japonicum | 門脈系統 |
棘口科 Echinostomatidae | 棘隙屬 Echinochasmus | 日本棘隙吸蟲 E. japonicus |