3 英文翻譯
Transcription factor
4 概述
在分子生物學中,轉錄因子(Transcription factor)是指能夠結合在某基因上游特異核苷酸序列上的蛋白質,這些蛋白質能調控其基因的轉錄。轉錄因子可以調控核糖核酸聚合酶(RNA聚合酶,或叫RNA合成酶)與DNA模板的結合。轉錄因子一般有不同的功能區域,如DNA結合結構域與效應結構域。轉錄因子不單與DNA序列上的啓動子結合,也可以和其它轉錄因子形成一轉錄因子聚合體來影響基因的轉錄。
基因轉錄有正調控和負調控之分。如細菌基因的負調控機制是當一種阻遏蛋白(repressor protein)結合在受調控的基因上時,基因不表達;而從靶基因上去除阻遏蛋白後,RNA聚合酶識別受調控基因的啓動子,使基因得以表達,這是正調控。這種阻遏蛋白是反式作用因子。
轉錄因子是起正調控作用的反式作用因子。轉錄因子是轉錄起始過程中RNA聚合酶所需的輔助因子。真核生物基因在無轉錄因子時處於不表達狀態,RNA聚合酶自身無法啓動基因轉錄,只有當轉錄因子(蛋白質)結合在其識別的DNA序列上後,基因纔開始表達。
轉錄因子的結合位點(transcription factor binding site,TFBS)是轉錄因子調節基因表達時,與mRNA結合的區域。按照常識,轉錄因子(transcription factor,TF)的結合位點一般應該分佈在基因的前端,但是,新的研究發現,人21和22號染色體上,只有22%的轉錄因子結合位點分佈在蛋白編碼基因的5'端。
真核生物在轉錄時往往需要多種蛋白質因子的協助。一種蛋白質是不是轉錄機構的一部分往往是通過體外系統看它是否是轉錄起始所必須的。一般可將這些轉錄所需的蛋白質分爲三大類:
(1)RNA聚合酶的亞基,它們是轉錄必須的,但並不對某一啓動子有特異性。
(2)某些轉錄因子能與RNA聚合酶結合形成起始複合物,但不組成遊離聚合酶的成分。這些因子可能是所有啓動子起始轉錄所必須的。但亦可能僅是譬如說轉錄終止所必須的。但是,在這一類因子中,要嚴格區分開哪些是RNA聚合酶的亞基,哪些僅是輔助因子,是很困難的。
(3)某些轉錄因子僅與其靶啓動子中的特異順序結合。如果這些順序存在於啓動子中,則這些順序因子是一般轉錄機構的一部分。如果這些順序僅存在於某些種類的啓動子中,則識別這些順序的因子也只是在這些特異啓動子上起始轉錄必須的。
5 真核生物轉錄過程機制
在真核細胞(eukaryotic cell)中,一個蛋白要由基因上的DNA模板(template)開始表現出來,必須經過許多調控的關卡(見圖一)。首先是轉錄作用(transcription):即以DNA分子爲模板,經RNA聚合酶作用合成信使RNA(messenger RNA, mRNA);然後經過包括剪接(splicing)、加蓋(capping)以及mRNA尾端聚腺嘌呤的添加(poly(A)tail adding)等處理控制(processing control)來修飾mRNA的結構和信息。然後再經由輸送控制(transport control)階段送到細胞質中特定的位置進行轉譯作用(translation):即按照mRNA的核 酸序列轉譯成蛋白的胺基酸序列。這些初步形成的蛋白,得再經過另外一些複雜的修飾(modification)才能成爲具有特定功能的蛋白。但其中mRNA及蛋白的穩定性〔由降解率(degradation rate)得知〕也影響到蛋白的表現量。目前,對於以上衆多的調控中,以合成mRNA的基本轉錄作用(basal transcription)機制之研究發展得最迅速,尤其是在這三到四年間突飛猛進。本文主要探討的就是mRNA由RNA聚合酶二型(RNA polymerase Ⅱ)所主導的基本轉錄作用之起始機制(initiation)。
 |
| 見圖一 |
5.1 基本轉錄與轉錄因子
所謂基本轉錄作用是指在沒有活化子(activator;可讓轉錄加速的蛋白)或抑制子(repressor;可讓轉錄減慢甚至停止的蛋白)的情況下所進行的基本轉錄過程。
基本轉錄作用的開始需要許多基本因子(basal factor),稱爲一般轉錄因子(general transcription factor, GTF),簡稱轉錄因子(transcription factor, TF),每個轉錄因子在轉錄起始作用時所扮演的角色都不盡相同。目前對這方面的研究如雨後春筍般地發展開來,因此所知道的也比較清楚了。
步驟一:TFⅡD
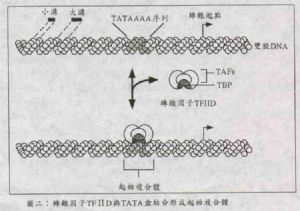
如圖二,一般DNA的轉錄起點前面都有一段DNA序列叫做啓動子(promoter),是由腺嘌呤(adenine, A)及胸腺嘧啶(thymine, T)所組成的,稱爲TATA盒(TATA box),其標準序列(consensus sequence)爲TATAAAA。
 |
| 圖二 |
在最先,轉錄因子TFⅡD蛋白(分子量120~140kDa)會找出這段TATA盒序列,並與這段DNA序列進行蛋白/DNA結合作用。早期的研究指出,TFⅡD是一個複合體(complex),由多個多胜肽鏈(polypeptide)組合而成,因此在TFⅡD複合體中專門與DNA結合的蛋白,我們就稱它爲TATA結合蛋白(TATA binding protein,簡稱TBP)。
TBP的分子量因不同的生物體而異,由27kDa(酵母菌)到38kDa(果蠅與人類細胞)都有。雖然TBP專責與DNA結合,但TBP跟其他能與DNA結合的DNA結合蛋白(DNA binding protein,簡稱DBP)有許多不同的地方。首先,TBP只需要單獨行動即可,是以單元體(monomer)形式和DNA作用,而非一般DBP要形成二聚體(dimer)才能和DNA結合(可進一步參考《科學月刊》25卷8期〈簡述視黃酸的訊息傳遞〉中以二聚體形式作用的DBP)。其次,TBP是與DNA分子螺旋結構上的小溝(minor groove)地方作用,而非一般DBP作用是在大溝(major groove)上;另外,TBP在蛋白上本身無法區分具有特定功能的功能部位(domain),亦即當TBP缺乏一些胺基酸片段後,它的DNA結合能力與轉錄啓動活性均告喪失(參考《科學月刊》25卷8期〈簡述視黃酸的訊息傳遞〉中有功能部位區分的核中受體)。從分子結構層次來看,TBP整個分子呈對稱的馬鞍狀,與DNA的結合方式就好像馬鞍套在馬背上一樣。當TBP與DNA結合後,除了TBP蛋白本身的形狀改變外,DNA亦會被扭曲,以便讓以後其他的轉錄因子來作用(可參閱1993年十月份英國的《Nature》中的許多計算機立體圖形)。
TFⅡD除了TBP之外,還有另外六個蛋白,它們被稱爲TBP附着因子(TBP-associated factor, TAF),能與其他TFⅡD以外的蛋白作用,例如活化子與抑制子,所以它們是負責轉錄活性調控的功能。
由於TFⅡD中的TBP結合到DNA的TATA序列上,是整個轉錄作用的首要之務,隨後引發一連串事件,最終促使轉錄之進行,因此這個蛋白(TFⅡD)與DNA的複合物,稱爲起始複合體(initiation complex)。
在早期的實驗指出,另外有一個轉錄因子TFⅡA會結合到起始複合體上並穩定其結構,但目前已證實TFⅡA只是在純化TFⅡD時一起沉澱下來的蛋白而巳,在轉錄的起始過程中並非是必需的因子。
步驟二:TFⅡB
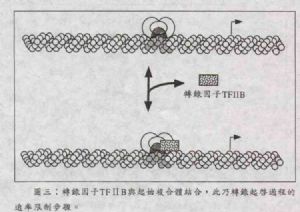
如圖三,轉錄因子TFⅡB對起始複合體的結合與否,爲轉錄起始作用過程的速率限制步驟(rate-limiting step)。而RNA聚合酶Ⅱ能否與起始複合體結合並開始轉錄作用,也完全視乎TFⅡB的結合與否。一些能使轉錄作用增強的活化子之所以能使轉錄作用加速,其原因就是可以加速TFⅡB結合到起始複合體上。
 |
| 圖三 |
TFⅡB爲單一蛋白結構,分子量由35kDa(老鼠細胞)到41kDa(酵母菌)都有。TFⅡB的功能是讓RNA聚合酶結合到起始複合體上,並且測定出轉錄起始點的位置。
步驟三:TFⅡF與RNA聚合酶
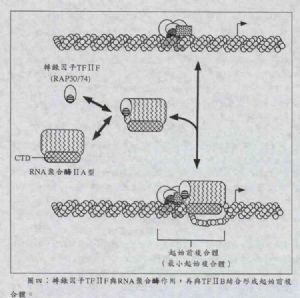
圖四中,另外一個轉錄因子TFⅡF是由兩個分子量分別爲30kDa與74kDa的多胜肽鏈所組成的異型二聚體(heterodimer),所以又稱爲RAP30/74。它首先在細胞核中與未和DNA結合的遊離RNA聚合酶結合,然後攜帶着RNA聚合酶與前述在起始複合體上的TFⅡB結合;因此TFⅡF就好像「媒人」一般,拖着RNA聚合酶與TFⅡB認識並與之結合。目前已知能與RNA聚合酶結合的蛋白除了TFⅡF與TFⅡB之外,前述TFⅡD中的TBP也被證實有跟RNA聚合酶結合的能力。
 |
| 圖四 |
值得一提的是:RNA聚合酶二型在轉錄過程中有兩個形式存在,一是沒有被磷酸化(non-phosphorylation)時稱爲A型(即RNA polymerase ⅡA),而如果已被磷酸化(phosphorylation),則稱爲O型(即RNA polymerase ⅡO)。磷酸化的位置集中在RNA聚合酶的羰基端,稱之爲羰基端功能部位(carboxyl-terminal domain, CTD)。而TFⅡF只與RNA聚合酶A型作用。
這時,RNA聚合酶A型結合到DNA上,整個複合體已有能力打開DNA的雙股螺旋結構了,但它還未有足夠的能力移動而停留在原地,這時的DNA與衆多的蛋白所形成的複合體,稱爲起始前複合體(pre-initiation complex, PIC)或最小起始複合體(minimal initiation complex)。
步驟四:TFⅡE
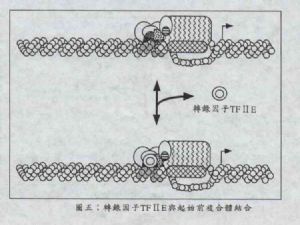
圖五中,轉錄因子TFⅡE已證實能與前述的TBP、TFⅡF及RNA聚合酶ⅡA型、和下一步的轉錄因子TFⅡH結合(詳見步驟五)。TFⅡE由兩個多胜肽鏈(分別爲34kDa及56kDa)所組成,它與PIC的結合可以穩定PIC,並引導下一個轉錄因子TFⅡH往PIC上結合。
 |
| 圖五 |
從一開始的第一步(TFⅡD與DNA上的TATA序列結合),一直至目前(TFⅡE與PIC結合),所有的蛋白與DNA或蛋白與蛋白的結合反應都是可逆的步驟(reversible pathway),所以表示轉錄因子就算結合後也不一定會往下一步走。
步驟五:TFⅡH
圖六中,當轉錄因子TFⅡH(又稱爲BTF2)結合到TFⅡE上時,整個包括RNA聚合酶的複合體可稱爲完整起始複合體(complete initiation complex)。TFⅡH是一個多功能蛋白,由五個多胜肽鏈所構成的複合體(大約有200kDa),它最特別的地方是:在目前衆多轉錄因子的功能中,只有它擁有激酶(kinase;進行磷酸化的酵素種類)、螺旋酶(helicase;能打開DNA螺旋結構的酵素)及ATP水解酶(ATPase;水解ATP分子而產生能量的酵素種類)的活性。而當TFⅡE及TFⅡH與PIC結合後,這時TFⅡH的ATP水解酶活性就會水解ATP分子而釋出能量,而螺旋酶的活性也可使雙股DNA打開的地方更大,這時期稱爲啓動子廓清(promoter clearance)階段,它可以說是介於啓動複合體與延長複合體(elongation complex)之間。而TF ⅡH的激酶活性是用來對RNA聚合酶的CTD作磷酸化作用的,藉以把RNA聚合酶ⅡA型轉變成ⅡO型,而CTD被磷酸化之後,RNA聚合酶隨即可以移動並開始轉錄作用。我們可以想象TFⅡH是一個開關,可以打開RNA聚合酶的動力機制,使之前進。
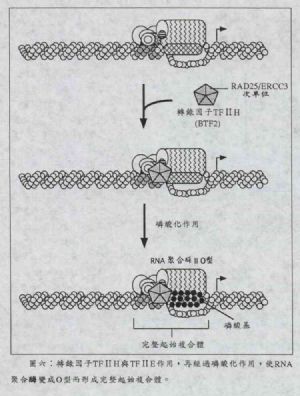 |
| 圖六 |
另外,在今年七月份英國《Nature》期刊中指出,TFⅡH中有一個與酵母菌RAD25蛋白很像的次單位(subunit),稱爲ERCC3(因此又稱爲RAD25/ERCC3次單位),它具有修補(repair)的作用,當轉錄中RNA聚合酶遇到有錯誤序列時,可能會促進修補機制之進行,而使轉錄機制與修補機制偶合在一起(transcription-repair coupling)。
步驟六:TFⅡG及TFⅡJ
目前還有兩個轉錄因子TFⅡG及TFⅡJ,但研究人員對其功能與存在的必要性仍有爭議,只知道它們出現在TFⅡH以後的步驟中。
步驟七:轉錄啓動
RNA聚合酶被磷酸化後,即可向前移動,當到達轉錄起啓點時,即可開始利用核 酸來進行聚合反應(polymerization)而合成RNA。
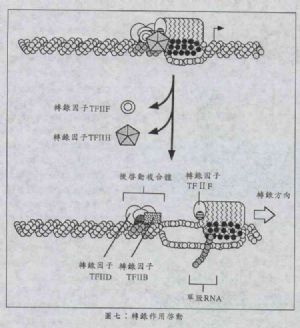
當RNA聚合酶向前移動時,完整起始複合體卻沒有完全分解開,反而分成三大陣營:一部分留在原地(TATA序列上),一部分跟着RNA聚合酶移動,剩下的部分解離出去。目前已知TFⅡD和TFⅡB停留不動,死咬着TATA序列不動;而TFⅡF與RNA聚合酶一起移動,共同進退;至於TFⅡE及TFⅡH則從複合體中解離出去,重獲自由身。
TFⅡD和TFⅡB停留在原地之複合物被稱爲後啓動複合物(post-initiation complex),它們賴着不走的目的主要是要與下一個轉錄因子作用,也就是說在下一輪的轉錄作用中,TFⅡD不必再花費時間和能量再去找TATA序列,TFⅡB也不必再重新找一遍TFⅡD,而重複做一次PIC的形成過程(見圖七)。
 |
| 圖七 |