2 英文參考
human leucocyte antigen,HLA
3 註解
排斥反應本質上是一種免疫反應,它是由組織表面的同種異型抗原誘導的。這種代表個體特異性的同種抗原稱爲組織兼容性抗原(histocompatibility antigen)或移植抗原(transplantation antigen)。機體內與排斥反應有關的抗原系統多達20種以上,其中能引起強而迅速排斥反應者稱爲主要組織兼容性抗原,其編碼基因是一組緊密連鎖的基因羣,稱爲主要組織兼容性複合體(major histocompatibilitycomplex,MHC)。控制機體免疫應答能力與調節功能的基因(immune uesponse gene,Ir gene )也存在於MHC內。因此,MHC不僅與移植排斥反應有關,也廣泛參與免疫應答的誘導與調節。人類的MHC稱爲人白細胞抗原系統(human leucocyte antigen,HLA)。
4 人類HLA基因複合體
對人主要組織兼容性抗原系統及其基因複合體的認識比小鼠約晚10年,法國學者Dausset在1958年首先發現,腎移植後出現排斥反應的患者以及多次輸血的患者血清中含有能與供者白細胞發生反應的抗體。後者所針對的抗原即人類主要組織相溶性抗原。由於該抗原首先在白細胞表面被發現且含量最高,而且白細胞抗原(human leucocyte antigen,HAL);人類MHC,即編碼HLA的基因羣自然數爲HAL複合體。
4.1 HLA複合體定位及結構
HLA複合體位於人第6號染色體的短臂上。該區DNA片段長度約3.5~4.0×千個鹼基對,占人體整個基因組的1/3000。圖5-2顯示HLA複合體結構。HLA複合體共有數十個座,傳統上按其產物的結構、表達方式、組織分佈與功能可將這些基因座分爲三類。
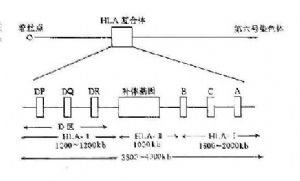
圖5-2 人類HLA複合體結構示意圖
1.HLA-Ⅰ類基因在Ⅰ基因區內存在多達31個有關的Ⅰ類基因座,其中HLA-A、HLA-B和HLA-C爲經典的HLA-Ⅰ類基因,其它基因的產物分佈有限,且其功能不明,另外還有許多僞基因.
2.HLA-Ⅱ類基因 HLA-Ⅱ類基因區包括近30個基因座,其中經典的Ⅱ類基因一般指DR、DP和DQ,它們編碼的產物均爲雙肽鏈(α、β)分子。近年來,陸續發現了一些位於Ⅱ類基因區的新基因座,其中某些基因的產物與內源性抗原的處理與呈遞有關。
3.HLA-Ⅲ類基因 HLA-Ⅲ類基因區域至少已發現36個基因座,其中C2、C4、Bf座編碼相應的補體成分,另外還有21羧化酶基因(CYP21A、B)腫瘤壞死因子基因(TNFA、B)以及熱休克蛋白70(heat shock protein70,HSP70)基因。補體C4由二個不同的基因(C4A與C4B)編碼。HLA-Ⅲ類基因區結構見圖5-3。

4.2 HLA等位基因及編碼產物的命名
按WHO-HLA命名委員會發布的資料,僅經典的HLA-Ⅰ、Ⅱ類座(A、B、C、DR、DQ、DP)等位基因即達279個。表5-1列出了至1991年11月已識別的HLA特異性。根據該委員會制定的命名原則,凡確定新的HLA抗原特異性都要明確其DNA序列。此外,下列幾種情況在HLA特異性編號後加W(work shop)標記。
表5-1 已識別的HLA特異性(1991)
| A | B | C | D | DR | DQ | DF |
| A1 | B5 B51(5) | Cw1 | Dw1 | DR1 | DQ1 | DPw1 |
| A2 | B7 B5102 | Cw2 | Dw2 | DR103 | DQ2 | DPw2 |
| A210(2) | B703 B5103(7) | Cw3 | Dw3 | DR2 | DQ3 | DPw3 |
| A3 | B8 B52(5) | Cw4 | Dw4 | DR3 | DQ4 | DPw4 |
| A9 | B12 B53 | Cw5 | Dw5 | DR4 | DQ5(1) | DPw6 |
| A10 | B13 B54(22) | Cw6 | Dw6 | DR5 | DQ6(1) | |
| A11 | B14 B55(22) | Cw7 | Dw7 | DR6 | DQ7 | |
| A19 | B15 B54(22) | Cw8 | Dw8 | DR7 | DQ18(3) | |
| A23(9) | B16 B57(17) | Cw9(w3) | Dw10 | DR8 | DQ19(3) | |
| A24(9) | B17 B54(17) | Cw1(w3) | Dw11(w7) | DR9 | ||
| A2403(9) | B18 B59 | Dw12 | DR10 | |||
| A25(10) | B21 B60(40) | Dw13 | DR11(5) | |||
| A26(10) | B22 B61(40) | Dw14 | DR12(5) | |||
| A28 | B27 B62(15) | Dw15 | DR13(5) | |||
| A29(19) | B35 B63(15) | Dw16 | DR13(6) | |||
| A30(19) | B37B64k(14) | Dw17(w7) | DR14(6) | |||
| A31(19) | B38(16 B65(14) | Dw18(w6) | DR1403 | |||
| A32(19) | B39(16) B67 | Dw19(w6) | DR1404 | |||
| A33(19) | B40 B70 | Dw20 | DR15(2) | |||
| A34(10) | B4005(21) B71(70) | Dw21 | DR16(2) | |||
| A36 | B41 B72(70) | Dw22 | DR17(3) | |||
| A43 | B42 B73 | Dw23 | DR18(3) | |||
| A66(10) | B44(12) B75(15) | DR51 | ||||
| A68(28) | B45(12) B76(15) | Dw24 | ||||
| A69(28) | B46 B77(15) | Dw25 | DR52 | |||
| A74(19) | B47 B7801 | B-26 | DR53 | |||
| B48 | ||||||
| B49(21) Bw4 | ||||||
| B50(21) Bw6 |
①Bw4和Bw6作爲表位以其它B座等位基因個區別;②C座的特異性加w,以與補體相區分;③由經典細胞學分型方法鑑定D和DP特異性加W。
4.3 HLA複合機遺傳特徵
1.單體型遺傳方式 HLA複合體是一組緊密連鎖的基因羣。這些連鎖在一條染色體上的等位基因很少發生同源染色體間的交換,構成一個單體型(haplotype)。在遺傳過程中,HLA單體型作爲一個完整的遺傳單位由親代傳給子代。有必要區分HLA表型、基因型與單體型這三個概念。某一個體HLA抗原特異性型別稱爲表型(phenotype);HLA基因在體細胞兩條染色體上的組合稱爲基因型(genotype);HLA基因在同一條染色體上的組合稱爲單體型(haplotype)(表5-2)。
| 受檢者 | 甲 A1 A2 B8 B12 | 乙 A1 A1 B8 B12 | 丙 A1 A1 B8 B8 |
| 表型 | HLA-A1、2:B8、12 | HLA-A1:B8、12 | HLA-A1、B8 |
| 基因型 | HLA-A1、A2 HLA-B8、B12 | HLA-A1、A1 HLA-B8、B12 | HLA-A1、A1 HLA-B8、B8 |
| 單體型 | HLA-A1、B8/A2、B12 | HLA-A1、B8/A1、B12 | HLA-A1、B8/A1、B8 |
二倍體(diploid)生物的每一細胞均有兩個同源染色體組,分別來自父母雙方。故子女的HLA單體型也是一個來自父方,一個來自母方。在同胞之間比較HLA單體型型別只會出現下列三種可能性:二個單體型完全相同或完全不同的機率各佔25%;有一個單體型相同的機率佔50%。至於親代與子代之間則必然有一個單體型相同,也只能有一個單體型相同(圖5-40。這一遺傳特點在器官移植供者的選擇以及法醫的親子鑑定中得到了應用。

注:a、b、c、d代表單體型
A1、B8、A2、B35等代表HLA基因座等位基因
2.多態性現象 多態性(polymorphism)是指在一隨機婚配的羣體中,染色體同一基因座有兩種以上基因型,即可能編碼二種以上的產物。HLA複合體是迄今已知人體最複雜的基因複合體,有高度的多態性。HLA的多態性現象乃由於下列原因所致:①復等位基因(multiple alleles):位於一對同源染色體上對應位置的一對基因稱爲等位基因(allele);由於羣體中的突變,同一座的基因系列稱爲復等位基因。前已述及,HLA複合體的每一座均存在爲數衆多的復等位基因,這是HLA高度多態性的最主要原因。由於各個座位基因是隨機組合的,故人羣中的基因型可達108之多。②共顯性(codominance);一對等位基因同爲顯性稱爲共顯性。HLA複合體中每一個等位基因均爲共顯性,從而大大增加了人羣中HLA表型的多樣性,達到107數量級。因此,除了同卵雙生外,無關個體間HLA型別全相同的可能性極小。
HLA的高度多態性顯示了遺傳背景的多樣性,這可能是高等動物抵禦不利環境因素的一種適應性表現,從而維持種屬的生存與延續具有重要的生物意義,但也對組織移植過程中尋找配型合的供體帶來很大的困難。
3.連鎖不平衡 HLA複合體各等位基因均有其各自的基因頻率。基因頻率是指某一特定等位基因與該基因座中全部等位基因總和的比例。隨機婚配的羣體中,在無新的突變和自然選擇的情況下,基因頻率可以代代維持不變,由於HLA複合體和各基因座是緊密連鎖的,若各座的等位基因隨機組合構成單體型,則某一單體型型別的出現頻率應等於該單體型各基因比其它基因能更多或更少地連鎖在起,從而出現連鎖不平衡(linkage disepuilibrium)。例如,在北歐白人中HLA-A1和HLA-B8頻率分別爲0.17和0.11。若隨機組合,則單體型A1-B8的預期頻率爲0.17×0.11=0.019。但實際所測行的A1-B8單體型頻率是0.088故A1-B8處於連鎖不平衡,實測頻率與預期頻率間的差值(△0.088-0.19=0.069)爲連鎖不平衡參數。在HLA複合體中已發現有50對以上等位基因顯示連鎖不平衡。產生連鎖不平衡的機制尚不清楚。
5 HLA抗原的分子結構
1987年Bjorkman等首先借助X線晶體衍射技術弄清了HLA-A2分子的立體結構。其後,其它HLA-Ⅰ、Ⅱ類分子結構的研究也取得了進展,從而對這些分子的生物學功能提供了較確切的解釋。
(一)HLA-Ⅰ類分子
所有的HLA-Ⅰ類分子均含有二條分離的多肽鏈,一條是由MHC基因編碼的α鏈或稱重鏈(44kD)。根據對HLA-A2和Aw68分子的晶體結構分析,Ⅰ類分子可分爲四個區:(圖5-5):①氨基端胞外多肽結合區:該區由二個相似的各包括90個氨基酸殘基的片段組成,分別稱爲α1和α2。該功能區含有與抗原結合的部位。後者呈深槽狀,其大小與形狀適合於已處理的抗原片段,約容納8~10個氨基酸殘基。Ⅰ類分子的多態性殘基也位於該區域。②胞外lg樣區:該區又稱爲重鏈的α3片段,包括90個氨基酸殘基,與免疫球蛋白的恆定區具有同源性。Ⅰ類分子與TC細胞表面CD8分子的結合部位即在α3片段。Ⅰ類分子的β鏈又稱β2微球蛋白,也結合於該區。β鏈由第15號染色體的基因編碼,它不插入細胞膜而遊離於細胞外。β2微球蛋白與α1、α2、α3片段的相互作用對維持Ⅰ類分子於然構型的穩定性及其分子表達有重要意義。③跨膜區:該區氨基酸殘基形成螺旋狀穿過漿膜的脂質雙層,將類分子錨定在膜上。④胞漿區:該區位於胞漿中,可能與細胞內外信息傳遞有關。
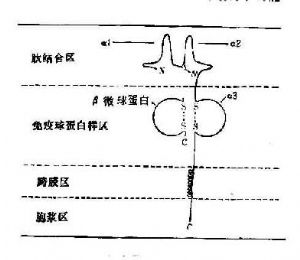
(二)HLA-Ⅱ類分子
所有的Ⅱ類分子均由二條以非共價鍵連接的多肽鏈(α、β)組成。二條鏈的基本結構相似,但分別由不同的MHC基因編碼,且均具有多態性。雖然Ⅱ類分子的晶體衍射結構尚未得到,但光譜分析已證明與Ⅰ類分子具有某種相似性。Ⅱ類分子二條多肽鏈也可分爲四個區,見(圖10-6):①肽結合區:α鏈與β鏈的胞外部位均可再分爲二個各含90個氨基酸殘基的片段,分別稱爲α1、α2和β1、β2。肽結合區包括α1和β1片段,該二片段構成肽結合的裂隙(cleft),約可容納14個氨基酸殘基。Ⅱ類分子的多態性殘基主要集中在α1和β1片段,這種多態性決定了多肽結合部位的生化結構,也決定了與肽類結合以及T細胞識別的特異性和親和力。②lg樣區:此區由α2和β2片段組成,兩者均含鏈內二硫鍵,並屬於lg基因超家族。在抗原呈遞過程中,TH細胞的CD4分子與Ⅱ類分子結合的部位即位於該lg樣非多肽態區域。③跨膜區和胞漿區:該二區與Ⅰ類分子α鏈的相應區域結構相似。
6 HLA抗原的組織分佈
各類HLA抗原的組織分佈不同。Ⅰ類抗原廣泛分佈於體內各種有核細胞表面,包括血小板和網織紅細胞。除某些特殊血型者外,成熟的紅細胞一般不表達Ⅰ類抗原。不同的組織細胞表達Ⅰ類抗原的密度各異。外周血白細胞和淋巴結、脾細胞所含Ⅰ類抗原量最多,其次爲肝、皮膚、主動脈和肌肉。但神經細胞和成熟的滋養層細胞不表達Ⅰ類抗原。Ⅱ類抗原主要表達在某些免疫細胞表面,如B細胞、單核/巨噬細胞,樹突狀細胞,激活的t 細胞等,內皮細胞和某些組織的上皮細胞也可檢出HLA-Ⅱ抗原。另外,某些組織細胞在病理情況下也可異常表達Ⅱ類抗原。Ⅰ、Ⅱ類抗原主要分佈在細胞表面,但也可能現於體液中,血清、尿液、唾液、精液及乳汁中均已檢出可溶性HLA-Ⅰ、Ⅱ類抗原。HLA-Ⅲ類抗原一般指幾種補體成分,它們均分佈於血清中。
7 HLA抗原表達的調控
在各類型細胞表面HLA分子表達與否以及表達的密度,可以受不同的因素調節。一般認爲,調控HLA分子表達的主要環節是轉錄速率。可能影響HLA分子表達的因素有:①組織細胞的分化階段:HLA分子是造血幹細胞和某些免疫細胞的分化抗原,在細胞分化、成熟的不同階段,各類HLA抗原的表達可有改變。例如HLA-DQ分子是人單核細胞的成熟標記;Ⅱ類抗原僅表達在激活的T細胞表面。②某些疾病狀態:某些傳染性疾病、免疫性疾病、造血系統疾病以及腫瘤均可影響HLA抗原表達。如AIDS病患者單核細胞HLA-Ⅱ類抗原表達明顯減少,某些腫瘤細胞表面HLA-Ⅰ類抗原表達減少。③生物活性物質:某些細胞因子,例如三類干擾素(α、β、γ)以及TNFα、THFβ均可增強不同類型細胞HLA-Ⅰ類抗原表達;具有Ⅱ類抗原誘生能力的細胞因子包括IFNγ、TNFα、IL-6及GM-CSF等。此外,某些激素、某些神經遞質和神經肽也可影響HLA分子表達。
HLA分子在免疫應答與免疫調節中是一類關鍵的分子,故各種因素對HLA分子表達的調控可能是體內免疫調節網絡的重要組成部分。同時,受各種調節因子的影響,HLA分子的異常表達也參與某些疾病的發病機制。
8 HLA與疾病相關性
不同個體對疾病易感性的差異在很大程度上是由遺傳因素所決定。在羣體調查中比較患者與正常人某些特定等位基因及其產物的頻率,這是研究遺傳決定的對疾病易感性的主要方法。
HLA是目前已知的具有最複雜多態性的人類基因系統,且Ir基因正位於HLA複合體內,因此考慮到HLA與某些免疫性疾病可能存在相關性。60年代末通過對患者與正常人HLA抗原頻率的羣體調查,發現了某些疾病與特定的HLA型分別呈非隨機分佈。最典型的例子是91%以上的北美白人強直性脊柱炎患者帶有HLA-B27抗原。這種現象,即二個遺傳學性狀在羣體中同時出現呈非隨機分佈,稱爲關聯(association)。HLA是第一個被發現與疾病有明確聯繫的遺傳系統。迄今已發現60餘種疾病與HLA有關聯,這些多屬於病因或發病機制未知、與免疫異常有關,或有家族傾向及環境誘發因素的疾病。特定疾病與某種HLA型別的相關性可通過相對危險性(relative risk,RR)來評估,其計算公式爲:
RR=P+×C―/P―×C+
式中P+爲具有某種抗原的病人數;C―爲不帶此抗原的對照組人數;P―爲不帶此抗原的病人數;C+爲具有此抗原的對照組人數。RR表示帶某種HLA抗原的人與無此種抗原的人在患某種疾病的危險性上的比值。RR=1時,兩者無關聯;若RR>4,則認爲此病與某種HLA抗原肯定有關聯;RR值越大,表示帶此抗原的人患某病的危險性越大。反之,若RR<1,表示帶此抗原者某病有抵抗性。
在評估HLA與疾病的相關性時須注意下面幾點:①發現HLA與某種疾病有關聯,並不意味着攜帶某抗原就一定會患某病,HLA本身並不是病因而僅僅是一種遺傳標誌;②HLA抗原在羣體中的分佈與民族、人種、地理環境等有關,在研究與疾病的關聯時應綜合分析纔有參考價值;③研究對象須是隨機選擇,無親緣關係的:對照組與疾病相關性可能有助於某些疾病的輔助診斷,疾病的預測、分類以及預後的判斷。表5-3列出某些疾病與HLA的相關性。
表5-3 HLA 和疾病的相關性
| 疾病 | HLA抗原 | 相對危險性RR |
| 強直性脊柱炎 | Ⅰ類型 B27 | >100 |
| 青少年類風溼性關節炎 | B27 | 24 |
| Reiter病 | B27 | 30~50 |
| 牛皮癬性關節炎 | B17 | 6 |
| Bechat綜合徵 | Cw6 | 9 |
| 發作性睡眠 | B51 | 10~15 |
| 尋常天皰瘡 | Ⅱ類 DR2 | 20 |
| I型糖尿病 | DR4 | 24 |
| 多發性硬化症 | DR3/DR4 | 20 |
| 全身性紅斑狼瘡 | DR2 | 4 |
| 全身性硬化症 | Ⅲ類 C4AQO C4BQO C4AQO | 6 11 9 |
迄今已檢出了衆多的HLA基因多態性標誌。因此,有可能在DNA水平上探討HLA與疾病的相關性,甚至發現一些與經典HLA抗原未表現出關聯,但與HLA基因型別關聯的疾病。可以預期,隨着DNA水平的研究不斷深入,最終有可能在HLA複合體中發現某些疾病的易感基因,甚至測出這些基因的核苷酸序列。這將有助於闡明某些疾病的發病機制,並在此基礎上制訂全新的防治措施。
HLA與疾病關聯的機制尚未完全清楚,已提出的一些學說包括:①分子模擬學說:該學說認爲,由於HLA抗原本身與某種病原物質相似,機體或者不能對該病原物質產生有效的免疫應答,或者在對病原物的應答中發生交叉反應反而損害了自身組織;②受體學說:HLA抗原可能作爲外來病原物質的受體,兩者結合導致組織損傷;③免疫應答基因學說:人的HLA基因就是Ir基因,特定的Ⅱ類基因型可能導致特定的異常免疫應答,從而表現爲易感某種疾病;④連鎖不平衡學說:特定的HLA基因可能與某病的易感基因連鎖,HLA型別僅是一種檢出的遺傳標誌。一般認爲,與HLA有關聯的不同病種可能有不同的機制。
9 HLA表達異常與疾病的關係
HLA表達異常即細胞表面HLA分子質與量的異常,可參與疾病發生。
9.1 HLA-Ⅰ類抗原表達異常
在小鼠及許多人類腫瘤或腫瘤衍生的細胞株均已發現MHC-Ⅰ類抗原表達缺失或密度降低。若將Ⅰ類基因轉染給腫瘤細胞株,則惡變細胞可發生逆轉,且浸潤性與轉移性消失或降低。這可能是由於MHC-Ⅰ類抗原缺失的腫瘤細胞不能補TC識別並攻擊,從而導致腫瘤免疫逃逸(sneaking through)。
9.2 HLA-Ⅱ類抗原表達異常
器官特異性自身免疫疾病的靶細胞可異常表達HLA-Ⅱ類抗原。諸如Graves病患者的甲狀腺上皮細胞、原發性膽管肝硬化患者的膽管上皮細胞、Ⅰ型糖尿病患者的胰島β細胞等均可發現HLA-Ⅱ抗原異常表達。其機制可能是局部感染誘生IFN-γ,後者誘導Ⅱ類抗原表達。Ⅱ類抗原乃抗原呈遞的效應分子,一旦靶細胞異常表達Ⅱ類抗原,就可能以組織特異性方式把自身抗原呈遞給自身反應性T細胞,從而啓動自身免疫反應。激活的自身反應性TH又可分泌大量IFN-γ,誘導更多的靶細胞表達Ⅱ類抗原,加重和延續自身免疫反應,最終導致遷延不愈的自身組織損傷。
10 HLA與排斥反應
移植物存活率很大程度上取決於供者和受者之間HLA型別相合的程度。在腎移植中,各HLA座配合的重要性依次爲HLA-DR、HLA-B、HLA-A。近年來特別重視HLA-DP對移植器官長期存活的意義。在骨髓移植中,爲預防嚴重的移植物抗宿主反應(graft versus host reaction,GVHR),一般要求從同胞中選擇HLA全相同的個體作爲供者。此外,某些輸血反應以及習慣性流產也與HLA不兼容所導致的排斥反應有關。
11 HLA與法醫
由於HLA複合體的高度多態性,在無關個體間HLA表型全相同的機率極低,故HLA複合體被看作是伴隨個體終生的特異性遺傳標記。藉助HLA基因型和(或)表型檢測,可用於法醫上的個體識別。另外,由於HLA複合體具有高度多態性以及單倍型遺傳的特點,使HLA分型成爲鑑定親子關係的重要手段。
12 HLA分型技術
HLA分型並不只是一種應用性的臨牀檢測指標,免疫遺傳學研究的發展,很大程度上依賴於以分型爲主要手段的HLA多態性分析。60年代建立的並不斷完善的血清學及細胞學分型技術主要側重於分析HLA產物特異性;80年代起建立的DNA分型方法則側重於基因的分型。
12.1 血清學分型技術
12.1.1 HLA-Ⅰ類抗原的檢測
HLA-A、B、C抗原型別鑑定均藉助微量淋巴細胞毒試驗(microlymphocytotoxicitytest)或稱補體依賴的細胞毒試驗(complement dependent cytotoxicitytest)。原理爲取已知HLA抗血清加入待測外周血淋巴細胞,作用後加入免補體,充分作用後加入染料,在倒置顯微鏡下判斷結果,着染的細胞爲死亡細胞,表示待檢淋巴細胞表面具有已知抗血清所針對的抗原。標準抗原清取自多次經產婦或計劃免疫志願者。
12.1.2 HLA-DR、DQ抗原檢測
該二抗原分型方法同HLA-Ⅰ類抗原,但所用抗血清須經過血小板吸收以去除針對Ⅰ類抗原的抗體。另外,待測細胞須是經純化的B細胞。
血清學分型是一項古老的技術,雖然近年來已建立許多新的分型技術,但血清學方法目前仍是HLA分型的基礎。
12.2 細胞學分型技術
HLA-Dw特異性與HLA-DP特異性可分別通過純合分型細胞(homozygote typing cell,HTC)及預致敏淋巴細胞試驗(primed lymphocyte test,PLT)檢測。二種方法的基本原理均是判斷淋巴細胞在識別非已HLA抗原決定簇後發生的增殖反應。由於分型細胞來源困難以及操作手續繁瑣,細胞學分型技術下正逐漸淘汰。
12.3 HLA的DNA分型技術
上述傳統的HLA分型方法有許多不足之處,近年來國內外已將HLA分型技術由抗原水平發展到基因水平。
12.3.1 限制性片段長度多態性檢測技術
這是首先建立的對多態性進行檢測的DNA分析技術。個體間抗原特異性來自氨基酸順序的差別,後者由編碼基因的鹼基順序不同所決定。這種鹼基順序的差別造成限制性內切酶識位置及酶切位點數目的不同,從而產生數量和長度不一的DNA酶切片段。用特異性探針對整個基因組DNA酶切片段進行雜交,即可分析限制性長度片段多態性(restriction fragment lengthpolymorphism,RFLP)。一定的內切酶組合所得到的HLA-RFLP可以和傳統方法測定的HLA特異性型別相關。80年代末發展起來的PCR(polymerase chain reaction)技術已被用於RFLP分析,即用等位特異限制酶裂解PCR擴增的片段,然後再進行分析,從而大提高了靈敏度。
12.3.2 PCR/SSO技術
此法乃用人工合成的HLA型別特異的寡核苷酸序列作爲探針,與待檢細胞經PCR擴增的HLA基因片段雜交,從而確定HLA型別,PCR技術可將HLA複合體上指定基因片段特異性地擴增5~6個數量級;而專門設計的SSO(序列特異的寡核苷酸sequencedpecific oligonucleotide)探針又能探測出等位基因間1~2個核苷酸的差異,故PCR/SSO技術具有靈敏度、特異性強、需樣本量少等優點。
12.3.3 PCR/SSP技術
目前常規的HLA-DNA分型技術,包括上述的PCR/RFLP、PCR/SSO等,最終均需用標記的特性探針與擴增產物進行雜交,再分析結果。PCR/SSP方法用乃設計出一整套等位基因組特異性引物(sequence specific primer,SSP),藉助PCR技術獲得HLA型別特異的擴增產物,可通過電泳直接分析帶型決定HLA型別,從而大大簡化了實驗步驟。
由於傳統方法在Ⅱ類抗原分型方面困難較大,故上述幾種基因分析型方法目前主要用於Ⅱ類基因座。此外,目前已建立的HLA基因分型技術還包括PCR單鏈構像多態性分析(PCR-single strand conformational polymorphism,PCR-SSCP)和PCR 異源二聚體電泳多態即PCR指紋圖(PCr fingerprinting)分析。DNA分型技術的應用,使HLA型別分析達到了更精細的水平,並因此發現了更多的HLA多態性。HLA的DNA分型技術現已成爲血清學方法的競爭者,並可能在不久的將來完全取而代之。
HLA是目前所知人體最複雜的遺傳多態性系統。HLA研究涉及免疫學、生物學、遺傳學、分子生物學、醫學等多個學科,並已發展成爲一個獨立的學科分支。迄今HLA研究已達到相當深入的水平,並在諸多方面取得顯著進展,包括HLA複合體結構;HLA分子結構及其表達的調控;HLA分子功能,尤其是在抗原處理、呈遞及T細胞識別中的作用;HLA的DNA分型及多態性研究;HLA與疾病的關係;HLA與移植的關係等。HLA研究不僅使器官移植成爲一種極有價值的治療手段,並給基礎與臨牀免疫帶來了突破性進展。已經證實,HLA複合體中存在控制免疫應答的基因以及HLA參與約束免疫細胞間相互作用,這表示HLA涉及生命活動的各個水平與多個方面。可以預期,對HLA的研究將繼續成爲免疫遺傳學最活躍的部分;對HLA的應用將擴展到基礎、臨牀、預防醫學的各個領域。