3 概述
類病毒是能侵染某些高等植物的非細胞形態病原體。作用類似病毒,但沒有蛋白質外殼,是一類較小的核糖核酸(RNA)分子。1971年首先在患馬鈴薯紡錘塊莖病的病薯中發現。這種病原體對RNA酶極爲敏感而不被蛋白酶或脫氧核糖核酸(DNA)酶破壞,分子量約爲120千道爾頓。對它研究較多的第納(Theoder Diener)把這類感染因子叫做類病毒,以和病毒區別。在電子顯微鏡下把變性的馬鈴薯紡錘塊莖病類病毒(PSTV)和雙鏈DNA相比較,推測它是單鏈環形RNA分子。後來經過序列測定證實,PSTV是自身鹼基序列高度互補的環狀RNA,含359個核苷酸,其最可能的二級結構是由許多短雙鏈區和短不配對區交替排列形成的,可構成天然類病毒在電子顯微鏡下所顯示的棒狀結構。20世紀70年代還發現一些造成巨大經濟損失的植物病害是類病毒引起的,如椰子死亡病類病毒曾殺死菲律賓島上約3000萬株椰子樹。目前已發現近20種類病毒,其中有10幾種已測定了一級結構。它們都是高度自身互補的單鏈閉環RNA。此外,
絕大多數類病毒有共同的結構特徵,它們的棒狀結構中心有一個高度保守的區域。包括PSTV在內的幾種類病毒沒有翻譯起始密碼子和mRNA的其他調控序列,現認爲,類病毒不能編碼蛋白質。對類病毒的複製、致病機制和起源還不清楚。
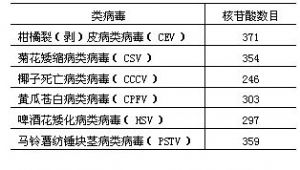
6 類病毒成員
| 科 | 屬 | 種 | 型 |
| 馬鈴薯紡錘形塊莖類病毒科(Pospiviroidae) | 馬鈴薯紡錘形塊莖類病毒屬 (Pospiviroid) | 菊矮化類病毒(Chrysanthemum stunt viroid) | 菊矮化類病毒-矮牽牛(Chrysanthemum stunt viroid-Petunia) |
| 柑橘裂皮類病毒(Citrus exocortis viroid) | 柑橘裂皮類病毒-蠶豆(Citrus exocortis viroid-Broad bean) | ||
| 柑橘裂皮類病毒-胡蘿蔔(Citrus exocortis viroid-Carrot) | |||
| 柑橘裂皮類病毒-茄子(Citrus exocortis viroid-Eggplant) | |||
| 柑橘裂皮類病毒-葡萄(Citrus exocortis viroid-Grapevine) | |||
| 柑橘裂皮類病毒-番茄(Citrus exocortis viroid-Tomato) | |||
| 柑橘裂皮類病毒-蕪菁(Citrus exocortis viroid-Turnip) | |||
| 金魚花潛隱類病毒(Columnea latent viroid) | 金魚花潛隱類病毒-鴛鴦葉草(Columnea latent viroid-Brunfelsia undulata) | ||
| 金魚花潛隱類病毒-Nemathantus(Columnea latent viroid-Nemathantus) | |||
| 血莧類病毒 1號(Iresine viroid 1) | |||
| 墨西哥心葉茄類病毒(Mexican pepita viroid) | |||
| 馬鈴薯紡錘形塊莖類病毒(Potato spindle tuber viroid) | 馬鈴薯紡錘形塊莖類病毒-鱷梨(Potato spindle tuber viroid-avacado) | ||
| 馬鈴薯紡錘形塊莖類病毒-鳳果(Potato spindle tuber viroid-pepino) | |||
| 馬鈴薯紡錘形塊莖類病毒-茄屬(Potato spindle tuber viroid-solanum spp) | |||
| 番茄頂縮類病毒(Tomato apical stunt viroid) | 番茄頂縮類病毒-內辣茄(Tomato apical stunt viroid-Solanum pseudocapsicum) | ||
| 番茄萎黃矮化類病毒(Tomato chlorotic dwarf viroid) | |||
| 番茄整株結節類病毒(Tomato planta macho viriod) | |||
| 啤酒花矮化類病毒屬(Hostuviroid) | 啤酒花矮化類病毒(Hop stunt viroid | 啤酒花矮化類病毒-杏樹(Hop stunt viroid-almond) | |
| 啤酒花矮化類病毒-杏(Hop stunt viroid-apricot) | |||
| 啤酒花矮化類病毒-柑桔(Hop stunt viroid-citrus) | |||
| 啤酒花矮化類病毒-黃瓜(Hop stunt viroid-cucumber) | |||
| 啤酒花矮化類病毒-葡萄(Hop stunt viroid-grapevine) | |||
| 啤酒花矮化類病毒-桃子(Hop stunt viroid-peach) | |||
| 啤酒花矮化類病毒-梨(Hop stunt viroid-pear) | |||
| 啤酒花矮化類病毒-李(Hop stunt viroid-plum) | |||
| 椰子死亡類病毒屬 (Cocadviroid) | 柑橘類病毒IV 號(Citrus viroid IV) | ||
| 椰子死亡類病毒(Coconut cadang-cadang viroid) | |||
| 椰子敗生類病毒(Coconut tinangaja viroid) | |||
| 啤酒花潛隱類病毒(Hop latent viroid) | |||
| 蘋果鏽果類病毒屬 (Apscaviroid) | 蘋果凹果類病毒(Apple dimple fruit viroid) | ||
| 蘋果鏽果類病毒(Apple scar skin viroid) | 蘋果鏽果類病毒-斑點(Apple scar skin viroid-dapple) | ||
| 蘋果鏽果類病毒-日本梨(Apple scar skin viroid-Japanese pear) | |||
| 蘋果鏽果類病毒-梨鏽皮(Apple scar skin viroid-pear rusty skin) | |||
| 澳洲葡萄類病毒(Australian grapevine viroid) | |||
| 柑橘曲葉類病毒(Citrus bent leaf viroid) | |||
| 柑橘類病毒III號(Citrus viroid III) | |||
| 葡萄黃點類病毒1號(Grapevine yellow speckle viroid 1) | |||
| 葡萄黃點類病毒2號(Grapevine yellow speckle viroid 2) | |||
| 梨皰狀潰瘍類病毒(Pear blister canker viroid) | |||
| 蘋果果實皺紋類病毒(Apple fruit crinkle viroid) | |||
| 柑桔類病毒原始來源(Citrus viroid original source) | |||
| 柑桔類病毒V(Citrus viroid V) | |||
| 柑桔類病毒VI(Citrus viroid VI) | |||
| 錦紫蘇類病毒屬(Coleviroid) | 錦紫蘇類病毒1號(Coleus blumei viroid 1) | ||
| 錦紫蘇類病毒2號(Coleus blumei viroid 2) | |||
| 錦紫蘇類病毒3號(Coleus blumei viroid 3) | |||
| 鱷梨日斑類病毒科(Avsunviroidae) | 鱷梨日斑類病毒屬(Avsunviroid) | 鱷梨日斑類病毒(Avocado sunblotch viroid) | |
| 桃潛隱花葉類病毒屬(Pelamoviroid) | 菊花褪綠班駁類病毒(Chrysanthemum chlorotic mottle viroid) | ||
| 桃潛隱花葉類病毒(Peach latent mosaic viroid) | 桃潛隱花葉類病毒-蘋果(Peach latent mosaic viroid-apple) | ||
| 桃潛隱花葉類病毒-杏(Peach latent mosaic viroid-apricot) | |||
| 桃潛隱花葉類病毒-櫻桃(Peach latent mosaic viroid-cherry) | |||
| 桃潛隱花葉類病毒-梨(Peach latent mosaic viroid-pear) | |||
| 桃潛隱花葉類病毒-李(Peach latent mosaic viroid-plum) | |||
| - | - | 烏飯樹花葉類病毒(Blueberry mosaic viroid-like RNA) | |
| - | - | 牛旁矮化類病毒(Burdock stunt viroid) | |
| - | - | 茄潛隱類病毒(Eggplant latent viroid) | |
| - | - | 心葉煙矮化類病毒(Nicotiana glutinosa stunt viroid) | |
| - | - | 木豆花葉斑駁類病毒(Pigeon pea mosaic mottle viroid) | |
| - | - | 番茄束頂類病毒(Tomato bunchy top viroid) | |
| - | 茄潛隱類病毒屬(Elaviroid) | 茄潛隱類病毒(Eggplant latent viroid) |
7 類病毒基本特性
類病毒是一類無衣殼蛋白、分子質量較低、能在寄主植物中自我複製的環狀單鏈RNA分子。分子量爲8×104~12.5×104,由246~399個核苷酸組成,G+C含量爲53%~60%,不編碼蛋白質,無抗原性。類病毒能耐受紫外線和作用於蛋白質的各種理化因素,比如對蛋白酶,胰蛋白酶,尿素等都不敏感(“真病毒”均敏感),在90℃下仍能存活(“真病毒”在50~60℃下失活)。
類病毒現在僅在高等植物中發現,大多數類病毒可分爲5個功能區,既左手末端區(TL),致病區(P),中央保守區(C),可變區(V)和右手末端區(TR)。TR和TL區的保守序列有利於複製酶的結合,與類病毒的複製起始有關。P區與類病毒對寄主植物的致病症狀有關。C區含95nt的中央保守序列,並有一個9nt的反向重複序列,能形成基環結構,它可能是類病毒複製中間體,可能是類病毒複製的一個重要控制區區域。V區的變異程度最大,即使是很接近的類病毒,同源性也小於50%,該區也與變異性有關。類病毒的複製與病毒有很大的不同,其RNA本身無mRNA活性,不編碼任何蛋白,因此其複製完全依賴於寄主的轉錄酶系統。所有類病毒的複製都是從RNA到RNA的直接轉錄,不涉及DNA,複製的最終產物是環狀類病毒(+)RNA分子。類病毒有時也以線狀小分子存在,這可能是尚未環化,或者是已環化的分子經核酶切割產生。複製按對稱和不對稱兩種滾環模式進行。對稱滾環有鱷梨日斑類病毒等,複製時先以(+)鏈爲模板轉錄生成多個單位長度的(-)鏈複製中間體,被切割成單位長度的線狀(-)鏈分子,自我環化後以環化的(-)鏈爲模板轉錄出多個單位長度的(+)鏈複製中間體,再進一步切割和連接,形成具有感染性的單體環化的類病毒分子。不對稱滾環模式有馬鈴薯紡錘形塊莖類病毒等,複製時以(+)鏈爲模板轉錄生成多個單位長度的(-)鏈複製中間體,不經剪切,直接作爲模板轉錄成(+)鏈複製中間體,再經過切割和連接,形成單體環化的類病毒分子。