3 HLA的基因組成
人類的主要組織相容性複合體(majorhistocompatibilitycomplex,MHC)稱爲HLA複合體,位於第6對染色體的短臂上,長度爲4分摩(centimorgan,cM),約爲4000kb。整個複合體上有近60個基因座,已正式命名的等位基因278個。根據編碼分子的特性不同,可將整個複合體的基因分成三類:Ⅰ類、Ⅱ類和Ⅲ類基因(圖6-2)。
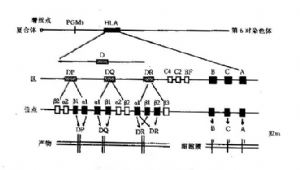
圖6-2HLA基因結構示意圖
1.類基因區位於着絲點的遠端,主要包括HLA-A、B、C三個位點;新近又提出E、F、G、H、K和L位點。
2.類基因區位於着絲點的近端,是結構最爲複雜的一個區,主要由DR、DQ、DP三個亞區構成,每個亞區又有若干個位點。新近又鑑定了DO、DZ、DX三個亞區。
3.類基因區含有編碼補體成分C2、C4、B因子及TNF、熱休克蛋白和21羥化酶的基因。
4.非HLA基因這些基因位於HLA區域內,其功能與HLA相關;目前已經命名的有兩類:LMP(largemultifunctionalprotease,或lowmolicularweightpolypeptides)和TAP(transporterassociatedwithantigenprocessing,或transporterofantigenpeptides)。LMP爲蛋白酶體相關基因,由LMP2和LMP7組成;TAP爲ABC轉運蛋白基因,包括TAP1和TAP2;它們的功能可能與抗原的處理和遞呈有關。
4 HLA的多態性
HLA複合體是人體最複雜的基因系統,呈高度的多態性,主要原因之一是由於HLA複合體的復等位基因所致。
遺傳學上將某一個體同源染色體上對應位置的一對基因稱爲等位基因(alleles);當羣體中位於同一位點的等位基因多於兩種時,稱爲復等位基因(muotiplealleles)。HLA複合體Ⅰ類和Ⅱ類基因位點多爲復等位基因。1995年公佈的用血清學、MLR和PLT確認的HLA特異性見表6-1。
| A | B | C | DR | DQ | DP | D | |
| A1 | B7 | B5102 | Cw1 | DR1 | DQ2 | DPw1 | Dw1 |
| A2 | B703 | B5103 | Cw2 | DR103 | DQ4 | DPw2 | Dw2 |
| A203 | B8 | B52(5) | Cw3 | DR2 | DQ5(1) | DPw3 | Dw3 |
| A210 | B13 | B53 | Cw4 | DR3 | DQ6(1) | DPw4 | Dw4 |
| A3 | B15 | B54(22) | Cw5 | DR4 | DQ793) | DPw5 | Dw5 |
| A11 | B18 | B55(22) | Cw6 | DR7 | DQ8(3) | DPw6 | Dw6 |
| A23(9) | B27 | B56(22) | Cw7 | DR8 | DQ9(3) | Dw7 | |
| A24(9) | B35 | B57(17) | Cw8 | DR9 | Dw8 | ||
| A2403 | B37 | B58(17) | Cw9(w3) | DR10 | Dw9 | ||
| A25(10) | B38(16) | B59 | DR11(5) | Dw10 | |||
| A26(10) | B39(16) | B60(40) | DR12(5) | Dw11(w7) | |||
| A29(19) | B3901 | B61(40) | DR13(6) | Dw12 | |||
| A30(19) | B3902 | B62(15) | DR14(6) | Dw13 | |||
| A31(19) | B40 | B63(15) | DR1403 | Dw14 | |||
| A32(19) | B4005 | B64(14) | DR1404 | Dw15 | |||
| A33(19) | B41 | B65(14) | DR15(2) | Dw16 | |||
| A34(10) | B42 | B67 | DR16(2) | Dw17(w7) | |||
| A36 | B44(12) | B70 | DR17(3) | Dw18(w6) | |||
| A43 | B45(12) | B71(70) | DR18(3) | Dw19(w6) | |||
| A66(10) | B46 | B72(70) | DR51 | Dw20 | |||
| A68(28) | B47 | B73 | DR52 | Dw21 | |||
| A69(28) | B48 | B75(15) | DR53 | Dw22 | |||
| A74(19) | B49(21) | B76(15) | Dw23 | ||||
| B50(21) | B77(15) | Dw24 | |||||
| B51(5) | B7801 | Dw25 | |||||
| Dw26 | |||||||
HLA抗原的命名由世界衛生組織命名委員會確定,每個特異性抗原均以其基因位點的字頭附以適當的數字(按抗原被發現或官方認可的順序)表示。標有w(workshop)的爲暫用名,得到認可後將其去掉;1991年決定:新特異性的申報要有明確的DNA順序,並根據DNA間關係命名,故取消w;現在所保留的w已非當初實驗室暫定名的含義,例如保留Cw以示與補體縮寫區別,保留Dw和DPw以示其用細胞學方法檢測。後面帶括弧的表示該特異性由括弧內的特異性分解而來,括弧內爲早期確認的抗原,包含多個特異性。表中D抗原不是獨立基因位點的編碼產物,而是與DR和DQ廣泛相關,是用細胞學方法檢測的抗原。
表6-1所列特異性是用血清學方法和細胞學方法鑑定出來的,幾乎每次會議都命名新的特異性。如此複雜的基因及產物,再加上單倍體共顯性遺傳的特點,可隨機組合成一個巨大的數字;以致在人羣中除同卵雙胎外,難以找到HLA完全相同者。這充分體現了HLA對免疫調控的個體差異,也爲同種器官移植增加了困難。
現在用分子生物學方法可在基因水平上鑑定出更大的HLA多態性,例如HLA-A2的基因有12個變異體(A*0201~A*0212),其差別僅在第19密碼子一個鹼基的置換。1994年3月WHO命名委員會公佈的Ⅰ類和Ⅱ類等位基因爲440個,1995年1月又發現了35個新的基因序列,並對以前的報告進行了部分修正。
5 HLA的遺傳特點
1.單倍型遺傳單倍型(haplotype)是指一條染色體上HLA各位點基因緊密連鎖組成的基因單位。人體細胞爲二倍體型,兩個單倍型分別來自父親和母親,共同組成個體的基因型(genotype)。由於一條染色體上HLA各位點的距離非常近,很少發生同源染色體之間的交換,因此新代的HLA以單倍型爲單位將遺傳信息傳給子代。例如父親的基因型爲ab,母親的爲cd,則子代可能有4種基因型,ac,ad,bc,bd,某一個體獲得任一基因型的可能性都是1/4。故兩個同胞有完全相同或完全不同HLA基因型的可能性都是1/4;一個單倍型相同的可能性是1/2。而子代和親代總是共有一個相同的單倍型。
2.共顯性遺傳共顯性(co-dominance)是指某位點的等位基因不論是雜合子還是純合子,均能同等表達,兩者的編碼產物都可在細胞表面檢測到。故每個位點可具有兩個抗原,可能相同,也可能不相同;這些抗原組成了個體的表型(phenotype)。多數個體的HLA位點都是雜合子,但當父親和母親在某位點上具有相同的等位基因時,其子代的這個位點就成爲純合子。
3.連鎖不平衡理論上,一個HLA位點的等位基因與另一個或幾個位點的等位基因在某一單倍型出現的頻率應等於各自頻率的乘積。然而在很多情況下,預期的單倍型頻率往往與實際檢測的頻率相差很大,在不同的地區或不同的人羣,某些基因相伴出現的頻率特別高,這種現象稱爲連鎖不平衡(linkagedisequilibrium)。HLA基因連鎖不平衡的發生機制目前尚不清楚,但已經發現某些疾病的發生與HLA複合體中某些特定的等位基因密切相關;某些連鎖不平衡傾向於出現在某些區域、某些人種和某些民族。深入探討連鎖不平衡的發生機制無疑將有助於對某些疾病的診斷和治療,亦將爲人類學研究增添新的內容。