2 註解
脫氧核糖核酸(英語:Deoxyribonucleic acid,縮寫爲DNA)又稱去氧核糖核酸,是一種分子,可組成遺傳指令,以引導生物發育與生命機能運作。主要功能是長期性的資訊儲存,可比喻爲“藍圖”或“食譜”。其中包含的指令,是建構細胞內其他的化合物,如蛋白質與RNA所需。帶有遺傳訊息的DNA片段稱爲基因,其他的DNA序列,有些直接以自身構造發揮作用,有些則參與調控遺傳訊息的表現。
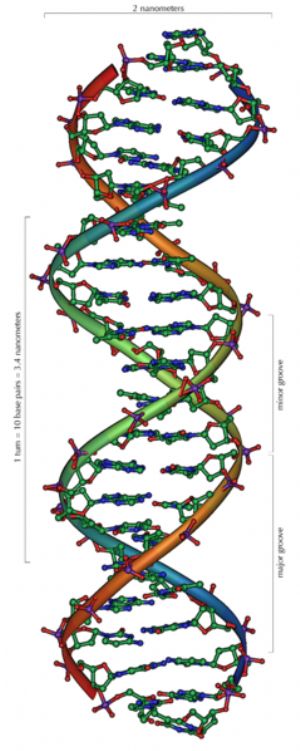 |
| DNA雙股螺旋。 |
DNA是一種長鏈聚合物,組成單位稱爲核苷酸,而糖類與磷酸分子藉由酯鍵相連,組成其長鏈骨架。每個糖分子都與四種鹼基裏的其中一種相接,這些鹼基沿着DNA長鏈所排列而成的序列,可組成遺傳密碼,是蛋白質氨基酸序列合成的依據。讀取密碼的過程稱爲轉錄,是根據DNA序列複製出一段稱爲RNA的核酸分子。多數RNA帶有合成蛋白質的訊息,另有一些本身就擁有特殊功能,例如rRNA、snRNA與siRNA。
在細胞內,DNA能組織成染色體結構,整組染色體則統稱爲基因組。染色體在細胞分裂之前會先行複製,此過程稱爲DNA複製。對真核生物,如動物、植物及真菌而言,染色體是存放於細胞核內;對於原核生物而言,如細菌,則是存放在細胞質中的類核裏。染色體上的染色質蛋白,如組織蛋白,能夠將DNA組織並壓縮,以幫助DNA與其他蛋白質進行交互作用,進而調節基因的轉錄。
3 歷史

佛朗西斯·克里克所繪,最早的DNA雙螺旋草圖。 參見:分子生物學史
最早分離出DNA的弗雷德里希·米歇爾是一名瑞士醫生,他在1869年,從廢棄繃帶裏所殘留的膿液中,發現一些只有顯微鏡可觀察的物質。由於這些物質位於細胞核中,因此米歇爾稱之爲“核素”(nuclein)。到了1919年,菲巴斯·利文進一步辨識出組成DNA的鹼基、糖類以及磷酸核苷酸單元,他認爲DNA可能是許多核苷酸經由磷酸基團的聯結,而串聯在一起。不過他所提出概念中,DNA長鏈較短,且其中的鹼基是以固定順序重複排列。1937年,威廉·阿斯特伯裏完成了第一張X光衍射圖,闡明瞭DNA結構的規律性。
1928年,弗雷德裏克·格里菲斯從格里菲斯實驗中發現,平滑型的肺炎球菌,能轉變成爲粗糙型的同種細菌,方法是將已死的平滑型與粗糙型活體混合在一起。這種現象稱爲“轉型”。但造成此現象的因子,也就是DNA,是直到1943年,才由奧斯瓦爾德·埃弗裏等人所辨識出來。1953年,阿弗雷德·赫希與瑪莎·蔡斯確認了DNA的遺傳功能,他們在赫希-蔡斯實驗中發現,DNA是T2噬菌體的遺傳物質。

劍橋大學裏一面紀念克里克與DNA結構的彩繪窗。
到了1953年,當時在卡文迪許實驗室的詹姆斯·沃森與佛朗西斯·克里克,依據倫敦國王學院的羅莎琳·富蘭克林所拍攝的X光衍射圖及相關資料,提出了最早的DNA結構精確模型,並發表於《自然》期刊。五篇關於此模型的實驗證據論文,也同時以同一主題發表於《自然》。其中包括富蘭克林與雷蒙·葛斯林的論文,此文所附帶的X光衍射圖,是沃森與克里克闡明DNA結構的關鍵證據。此外莫里斯·威爾金斯團隊也是同期論文的發表者之一。富蘭克林與葛斯林隨後又提出了A型與B型DNA雙螺旋結構之間的差異。1962年,沃森、克里克以及威爾金斯共同獲得了諾貝爾生理學或醫學獎。
克里克在1957年的一場演說中,提出了分子生物學的中心法則,預測了DNA、RNA以及蛋白質之間的關係,並闡述了“轉接子假說”(即後來的tRNA)。1958年,馬修·梅瑟生與富蘭克林·史達在梅瑟生-史達實驗中,確認了DNA的複製機制。後來克里克團隊的研究顯示,遺傳密碼是由三個鹼基以不重複的方式所組成,稱爲密碼子。這些密碼子所構成的遺傳密碼,最後是由哈爾·葛賓·科拉納、羅伯特·W·霍利以及馬歇爾·沃倫·尼倫伯格解出。爲了測出所有人類的DNA序列,人類基因組計劃於1990年代展開。到了2001年,多國合作的國際團隊與私人企業塞雷拉基因組公司,分別將人類基因組序列草圖發表於《自然》與《科學》兩份期刊。
4 物理與化學性質
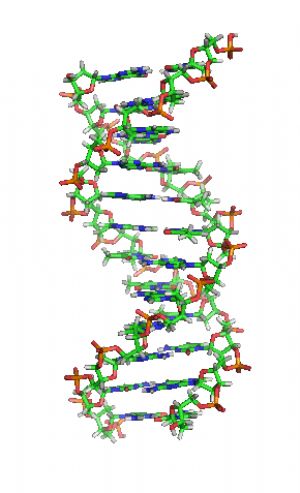
DNA片段結構動畫,各種鹼基水平排列於兩條螺旋長鏈之間。放大觀看
兩股DNA長鏈會以右旋方式相互纏繞成雙螺旋結構,由於以磷酸聯結而成的骨架位於外部,且兩股之間會留下一些空隙,因此位於螺旋內部的鹼基,即使從螺旋外側依然可見(如右方動畫)。雙螺旋的表面有兩種凹槽(或稱“溝”):較大的寬22埃;較小的寬12埃。由於各個鹼基靠近大凹槽的一面較容易與外界接觸,因此如轉錄因子等能夠與特定序列結合的蛋白質與鹼基接觸時,通常是作用在靠近大凹槽的一面。
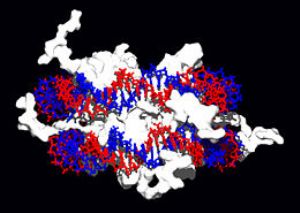
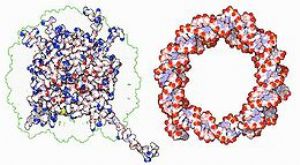
DNA與組織蛋白(上圖白色部分)的交互作用,這種蛋白質中的鹼性氨基酸(左下藍色),可與DNA上的酸性磷酸基團結合(右下紅色)。
結構蛋白可與DNA結合,是非專一性DNA-蛋白質交互作用的常見例子。染色體中的結構蛋白與DNA組合成複合物,使DNA組織成緊密結實的染色質構造。對真核生物來說,染色質是由DNA與一種稱爲組織蛋白的小型鹼性蛋白質所組合而成;而原核生物體內的此種結構,則摻雜了多種類型的蛋白質。雙股DNA可在組織蛋白的表面上附着並纏繞整整兩圈,以形成一種稱爲核小體的盤狀複合物。組織蛋白裏的鹼性殘基,與DNA上的酸性糖磷酸骨架之間可形成離子鍵,使兩者發生非專一性交互作用,也使複合物中的鹼基序列相互分離。在鹼性氨基酸殘基上所發生的化學修飾有甲基化、磷酸化與乙酰化等,這些化學作用可使DNA與組織蛋白之間的作用強度發生變化,進而使DNA與轉錄因子接觸的難易度改變,影響轉錄作用的速率。其他位於染色體內的非專一性DNA結合蛋白,還包括一種能優先與DNA結合,並使其扭曲的高移動性羣蛋白。這類蛋白質可以改變核小體的排列方式,產生更復雜的染色質結構。
DNA結合蛋白中有一種專門與單股DNA結合的類型,稱爲單股DNA結合蛋白。人類的複製蛋白A是此類蛋白中獲得較多研究的成員,作用於多數與解開雙螺旋有關的過程,包括DNA複製、重組以及DNA修復。這類結合蛋白可固定單股DNA,使其變得較爲穩定,以避免形成莖環(stem-loop),或是因爲核酸酶的作用而水解。
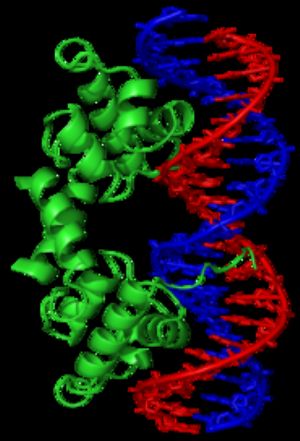
λ抑制子是一類具螺旋-轉角-螺旋結構的轉錄因子,可與DNA目標結合。
相對而言,其他的蛋白質則只能與特定的DNA序列進行專一性結合。大多數關於此類蛋白質的研究集中於各種可調控轉錄作用的轉錄因子。這類蛋白質中的每一種,都能與特定的DNA序列結合,進而活化或抑制位於啓動子附近序列的基因轉錄作用。轉錄因子有兩種作用方式,第一種可以直接或經由其他中介蛋白質的作用,而與負責轉錄的RNA聚合酶結合,再使聚合酶與啓動子結合,並開啓轉錄作用。第二種則與專門修飾組織蛋白的酵素結合於啓動子上,使DNA模板與聚合酶發生接觸的難度改變。
由於目標DNA可能散佈在生物體中的整個基因組中,因此改變一種轉錄因子的活性可能會影響許多基因的運作。這些轉錄因子也因此經常成爲信號傳遞過程中的作用目標,也就是作爲細胞反映環境改變,或是進行分化和發育時的媒介。具專一性的轉錄因子會與DNA發生交互作用,使DNA鹼基的周圍產生許多接觸點,讓其他蛋白質得以“讀取”這些DNA序列。多數的鹼基交互作用發生在大凹槽,也就是最容易從外界接觸鹼基的部位。
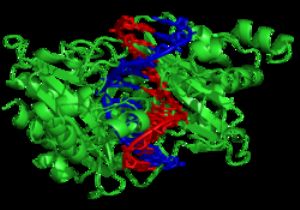
限制酶EcoRV(綠色)與其受質DNA形成複合物。
4.1 DNA修飾酵素
核酸酶與連接酶
核酸酶是一種可經由催化磷酸雙酯鍵的水解,而將DNA鏈切斷的酵素。其中一種稱爲外切酶,可水解位於DNA長鏈末端的核苷酸;另一種則是內切酶,作用於DNA兩個端點之間的位置。在分子生物學領域中使用頻率最高的核酸酶爲限制內切酶,可切割特定的DNA序列。例如左圖中的EcoRV可辨識出具有6個鹼基的5′-GAT|ATC-3′序列,並從GAT與ATC之間那條垂直線所在的位置將其切斷。此類酵素在自然界中能消化噬菌體DNA,以保護遭受噬菌體感染的細菌,此作用屬於限制修飾系統的一部分。在技術上,對序列具專一性的核酸酶可應用於分子選殖與DNA指紋分析。
另一種酵素DNA連接酶,則可利用來自腺苷三磷酸或煙酰胺腺嘌呤二核苷酸的能量,將斷裂的DNA長鏈重新接合。連接酶對於DNA複製過程中產生的延遲股而言尤其重要,這些位於複製叉上的短小片段,可在此酵素作用下黏合成爲DNA模板的完整複製品。此外連接酶也參與了DNA修復與遺傳重組作用。
拓撲異構酶與螺旋酶
拓撲異構酶是一種同時具有核酸酶與連接酶效用的酵素,可改變DNA的超螺旋程度。其中有些是先使DNA雙螺旋的其中一股切開以形成缺口,讓另一股能穿過此缺口,進而減低超螺旋程度,最後再將切開的部位黏合。其他類型則是將兩股DNA同時切開,使另一條雙股DNA得以通過此缺口,之後再將缺口黏合。拓撲異構酶參與了許多DNA相關作用,例如DNA複製與轉錄。
螺旋酶是分子馬達的一種類型,可利用來自各種核苷三磷酸,尤其是腺苷三磷酸的化學能量,破壞鹼基之間的氫鍵,使DNA雙螺旋解開成單股形式。此類酵素參與了大多數關於DNA的作用,且必須接觸鹼基才能發揮功用。
聚合酶
聚合酶是一種利用核苷三磷酸來合成聚合苷酸鏈的酵素,方法是將一個核苷酸連接到另一個核苷酸的3'羥基位置,因此所有的聚合酶都是以5'往3'的方向進行合成作用。在此類酵素的活化位置上,核苷三磷酸受質會與單股聚合苷酸模板發生鹼基配對,因而使聚合酶能夠精確地依據模板,合成出互補的另一股聚合苷酸。聚合酶可依據所能利用的模板類型來做分類。
在DNA複製過程中,依賴DNA模板的DNA聚合酶可合成出DNA序列的複製品。由於此複製過程的精確性是生命維持所必需,因此許多這類聚合酶擁有校正功能,可辨識出合成反應中偶然發生的配置錯誤,也就是一些無法與另一股配對的鹼基。檢測出錯誤之後,其3'到5'方向的外切酶活性會發生作用,並將錯誤的鹼基移除。大多數生物體內的DNA聚合酶,是以稱爲複製體的大型複合物形式來發生作用,此複合物中含有許多附加的次單位,如DNA夾或螺旋酶。
依賴RNA作爲模板的DNA聚合酶是一種較特別的聚合酶,可將RNA長鏈的序列複製成DNA版本。其中包括一種稱爲逆轉錄酶的病毒酵素,此種酵素參與了逆轉錄病毒對細胞的感染過程;另外還有複製端粒所需的端粒酶,本身結構中含有RNA模板。
轉錄作用是由依賴DNA作爲合成模板的RNA聚合酶來進行,此類酵素可將DNA長鏈上的序列複製成RNA版本。爲了起始一個基因的轉錄,RNA聚合酶會先與一段稱爲啓動子的DNA序列結合,並使兩股DNA分離,再將基因序列複製成信使RNA,直到到達能使轉錄結束的終止子序列爲止。如同人類體內依賴DNA模板的DNA聚合酶,負責轉錄人類基因組中大多數基因的RNA聚合酶II,也是大型蛋白質複合物的一部分,此複合物受到多重調控,也含有許多附加的次單位。
5 遺傳重組
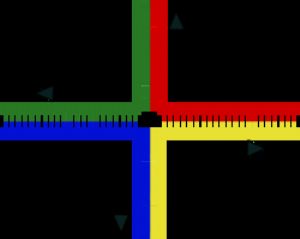
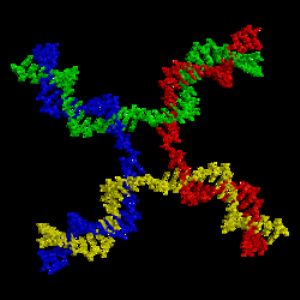
遺傳重組過程中產生的Holliday交叉結構,圖中的紅色、藍色、綠色與黃色分別表示四條不同的DNA長鏈。 參見:遺傳重組

重組過程中,兩條染色體(M與F)斷裂之後又重新接合,產生兩條重新排列過的染色體(C1與C2)。
各條DNA螺旋間的交互作用不常發生,在人類細胞核裏的每個染色體,各自擁有一塊稱作“染色體領域”的區域。染色體之間在物理上的分離,對於維持DNA資訊儲藏功能的穩定性而言相當重要。
不過染色體之間有時也會發生重組,在重組的過程中,會進行染色體互換:首先兩條DNA螺旋會先斷裂,之後交換其片段,最後再重新黏合。重組作用使染色體得以互相交換遺傳訊息,併產生新的基因組合,進而增加自然選擇的效果,且可能對蛋白質的演化產生重要影響。遺傳重組也參與DNA修復作用,尤其是當細胞中的DNA發生斷裂的時候。
同源重組是最常見的染色體互換方式,可發生於兩條序列相類似的染色體上。而非同源重組則對細胞具有傷害性,會造成染色體易位與遺傳異常。可催化重組反應的酵素,如RAD51,稱爲“重組酶”。重組作用的第一個步驟,是內切酶作用,或是DNA的損壞所造成的DNA雙股斷裂。重組酶可催化一系列步驟,使兩條螺旋結合產生Holliday交叉。其中每條螺旋中的單股DNA,皆與另一條螺旋上與之互補的DNA連結在一起,進而形成一種可於染色體內移動的交叉形構造,造成DNA鏈的互換。重組反應最後會因爲交叉結構的斷裂,以及DNA的重新黏合而停止。
6 DNA生物代謝的演化
DNA所包含的遺傳訊息,是所有現代生命機能,以及生物生長與繁殖的基礎。不過目前尚未明瞭在長達四十億年的生命史中,DNA究竟是何時出現並開始發生作用。有一些科學家認爲,早期的生命形態有可能是以RNA作爲遺傳物質。RNA可能在早期細胞代謝中扮演主要角色,一方面可傳遞遺傳訊息;另一方面也可作爲核糖酶的一部分,進行催化作用。在古代RNA世界裏,核酸同時具有催化與遺傳上的功能,而這些分子後來可能演化成爲目前以四種核苷酸組成遺傳密碼的形式,這是因爲當鹼基種類較少時,複製的精確性會增加;而鹼基種類較多時,增加的則是核酸的催化效能。兩種可達成不同目的功能最後在四種鹼基的情形下達到最合適數量。
不過關於這種古代遺傳系統並沒有直接證據,且由於DNA在環境中無法存留超過一百萬年,在溶液中又會逐漸降解成短小的片段,因此大多數化石中並無DNA可供研究。即使如此,仍有一些聲稱表示已經獲得更古老的DNA,其中一項研究表示,已從存活於2億5千萬年古老的鹽類晶體中的細菌分離出DNA,但此宣佈引起了討論與爭議。
7 技術應用
7.1 遺傳工程
重組DNA技術在現代生物學與生物化學中受到廣泛應用,所謂重組DNA,是指集合其他DNA序列所製成的人造DNA,可以質體或以病毒載體搭載所想要的格式,將DNA轉型到生物個體中。經過遺傳改造處裏之後的生物體,可用來生產重組蛋白質,以供醫學研究使用,或是於農業上栽種。
7.2 法醫鑑識
法醫可利用犯罪現場遺留的血液、精液、皮膚、唾液或毛髮中的DNA,來辨識可能的加害人。此過程稱爲遺傳指紋分析或DNA特徵測定,此分析方法比較不同人類個體中許多的重複DNA片段的長度,這些DNA片段包括短串聯重複序列與小衛星序列等,一般來說是最爲可靠的罪犯辨識技術。不過如果犯罪現場遭受多人的DNA污染,那麼將會變得較爲複雜難解。首先於1984年發展DNA特徵測定的人是一名英國遺傳學家阿萊克·傑弗里斯。到了1988年,英國的謀殺案嫌犯科林·皮奇福克,成爲第一位因DNA特徵測定證據而遭定罪者。利用特定類型犯罪者的DNA樣本,可建立出數據庫,幫助調查者解決一些只從現場採集到DNA樣本的舊案件。此外,DNA特徵測定也可用來辨識重大災害中的罹難者。
7.3 歷史學與人類學
由於DNA在經歷一段時間後會積聚一些具有遺傳能力突變,因此其中所包含的歷史訊息,可經由DNA序列的比較,使遺傳學家瞭解生物體的演化歷史,也就是種系。這些研究是種系發生學的一部分,也是演化生物學上的有利工具。假如對物種以內範圍的DNA序列進行比較,那麼羣體遺傳學家就可得知特定族羣的歷史。此方法的應用範圍可從生態遺傳學到人類學,舉例而言,DNA證據已被試圖用來尋找失蹤的以色列十支派。DNA也可以用來調查現代家族的親戚關係,例如建構莎麗·海明斯與托馬斯·傑斐遜的後代之間的家族關係,研究方式則與上述的犯罪調查相當類似,因此有時候某些犯罪調查案件之所以能解決,是因爲犯罪現場的DNA與犯罪者親屬的DNA相符。
7.4 生物資訊學
參見:生物資訊學
生物資訊學影響了DNA序列資料的運用、搜尋與資料挖掘工作,並發展出各種用於儲存並搜尋DNA序列的技術,可進一步應用於計算機科學,尤其是字串搜尋算法、機器學習以及數據庫理論。字串搜尋或比對算法是從較大的序列或較多的字母中,尋找單一序列或少數字母的出現位置,可發展用來搜尋特定的核苷酸序列。在其他如文本編輯器的應用裏,通常可用簡單的算法來解決問題,但只有少量可辨識特徵的DNA序列,卻造成這些算法的運作不良。序列比對則試圖辨識出同源序列,並定位出使這些序列產生差異的特定突變位置,其中的多重序列比對技術可用來研究種系發生關係及蛋白質的功能。由整個基因組所構成的資料含有的大量DNA序列,例如人類基因組計劃的研究對象。若要將每個染色體上的每個基因,以及負責調控基因的位置都標示出來,會相當困難。DNA序列上具有蛋白質或RNA編碼特徵的區域,可利用基因識別算法辨識出來,使研究者得以在進行實驗以前,就預測出生物體內可能表現出來的特殊基因產物。
7.5 DNA與電腦
參見:DNA運算
DNA最早在運算上應用,是解決了一個屬於NP完全的小型直接漢彌爾頓路徑問題。DNA可作爲“軟件”,將訊息寫成核苷酸序列;並以酵素或其他分子作爲“硬件”進行讀取或修飾。舉例來說,作爲硬件的限制酶FokI可以搭載一段具有軟件功能的GGATG序列DNA,再以其他的DNA片段進行輸入,並與軟硬件複合物產生反應,最後輸出另一段DNA。這種類似圖靈機的裝置可應用於藥物治療。此外DNA運算在能源消耗、空間需求以及效率上優於電子電腦,且DNA運算爲具有高度平行(見平行運算)的計算方式。許多其他問題,包括多種抽象機器的模擬、布爾可滿足性問題,以及有界形式的旅行推銷員問題,皆曾利用DNA運算做過分析。由於小巧緊密的特性,DNA也成爲密碼學理論的一部分,尤其在於能夠利用DNA有效地建構並使用無法破解的一次性密碼本。
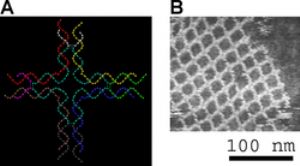
自我組裝產生的DNA納米結構。左方爲電腦繪圖,可見4條由DNA雙螺旋產生的交叉。右方爲原子力顯微鏡測得的影像。
7.6 DNA與納米科技
參見:DNA納米科技
DNA的分子性質,例如自我組裝特性,使其可用於某些納米尺度的建構技術,例如利用DNA作爲模板,可導引半導體晶體的生長。或是利用DNA本身,來製成一些特殊結構,例如由DNA長鏈交叉形成的DNA“瓦片”(tile)或是多面體。此外也可以做出一些可活動的元件,例如納米機械開關,此機械可經由使DNA在不同的光學異構物(B型與Z型)之間進行轉變,而使構形發生變化,導致開關的開啓或關閉。還有一種DNA機械含有類似鑷子的構造,可加入外來DNA使鑷子開合,並排出廢物DNA,此時DNA的作用類似“燃料”。DNA所建構出來的裝置,也可用來作爲上述的DNA運算工具。
9 延伸閱讀
- (中文)林正焜、洪火樹《認識DNA:下一波的醫療革命》。商周出版,2005年。ISBN 986-124-428-X
- (中文)Karl Drlica。周業仁譯。《DNA的14堂課》(Understanding DNA and Gene Cloning)。天下文化,2002年。ISBN 986-417-077-5
- (中文)James D. Watson。陳正萱、張項譯。《雙螺旋─DNA結構發現者的青春告白》(The Double Helix)。時報出版,1998年。ISBN 957-132-617-8
- (英文)Clayton, Julie. (Ed.).50 Years of DNA, Palgrave MacMillan Press, 2003. ISBN 978-1-40-391479-8
- (英文)Judson, Horace Freeland.The Eighth Day of Creation: Makers of the Revolution in Biology, Cold Spring Harbor Laboratory Press, 1996. ISBN 978-0-87-969478-4
- (英文)Olby, Robert.The Path to The Double Helix: Discovery of DNA, first published in October 1974 by MacMillan, with foreword by Francis Crick; ISBN 978-0-48-668117-7; the definitive DNA textbook, revised in 1994, with a 9 page postscript.
- (英文)Ridley, Matt.Francis Crick: Discoverer of the Genetic Code (Eminent Lives)HarperCollins Publishers; 192 pp, ISBN 978-0-06-082333-7 2006
- (英文)Rose, Steven.The Chemistry of Life, Penguin, ISBN 978-0-14-027273-4.
- (英文)Watson, James D. and Francis H.C. Crick. A structure for Deoxyribose Nucleic Acid (PDF).Nature171, 737–738, 25 April 1953.
- (英文)Watson, James D.DNA: The Secret of LifeISBN 978-0-375-41546-3.
- (英文)Watson, James D.The Double Helix: A Personal Account of the Discovery of the Structure of DNA (Norton Critical Editions). ISBN 978-0-393-95075-5
- (英文)Watson, James D. "Avoid boring people and other lessons from a life in science" New York: Random House. ISBN 978-0-375-421844
- (英文)Calladine, Chris R.; Drew, Horace R.; Luisi, Ben F. and Travers, Andrew A.Understanding DNA, Elsevier Academic Press, 2003. ISBN 978-0-12155089-9
10 外部鏈接
- (中文)核酸- 生物化學教學網頁
- (中文)畫說DNA-DNA from the beginning中文版,以動畫介紹DNA
- (英文)DNA Interactive- Flash交互式介紹
- (英文)DNA from the beginning
- (英文)The Register of Francis Crick Personal Papers- 佛朗西斯·克里克的文章
- (英文)Double Helix 1953–2003英國國家生物技術教育中心
- (英文)Double helix: 50 years of DNA,Nature
- (英文)Rosalind Franklin's contributions to the study of DNA
- (英文)U.S. National DNA Day- DNA日
- (英文)Genetic Education Modules for Teachers-DNA from the Beginning學習指導
- (英文)Listen to Francis Crick and James Watson talking on the BBC in 1962, 1972, and 1974
- (英文)DNA under electron microscope
- (英文)DNA Articles- 論文與訊息收集
- (英文)DNA coiling to form chromosomes
- (英文)DISPLAR: DNA binding site prediction on protein
- (英文)Dolan DNA Learning Center
- (英文)Olby, R. (2003)"Quiet debut for the double helix"Nature421(January 23): 402–405.
- (英文)Basic animated guide to DNA cloning
- (英文)DNA the Double Helix Game- 諾貝爾獎官方網站